La trehalosa en los alimentos aumenta la virulencia del clostridium difficile epidémico
Artículo comentado - adulto
Por el Pr. Harry Sokol
Gastroenterología y nutrición, Hospital Saint-Antoine, París, Francia
Espacio para el público general
Encuentra aquí tu espacio dedicado
Acerca de este artículo
Autor

Fecha de publicación
26 Agosto 2021
Fecha de actualización
19 Agosto 2024
Comentario al artículo original de Collins et al. (Nature 2018)
La incidencia de la infección con Clostridium difficile ha aumentado recientemente. Esta bacteria se ha convertido en un patógeno nosocomial dominante en América del Norte y en Europa, si bien se sabe poco sobre lo que desencadenó su aparición. Aquí, los autores muestran que dos ribotipos epidémicos (RT027 y RT078) han adquirido mecanismos únicos para metabolizar bajas concentraciones del disacárido trehalosa que se utiliza como aditivo alimentario. El ribotipo 027 (RT027) contiene una sola mutación puntual en el represor transcripcional* de la trehalosa que aumenta su sensibilidad a la sustancia más de 500 veces. Además, la trehalosa en los alimentos aumenta la virulencia de las cepas RT027 en un modelo murino de infección con Clostridium difficile . El ribotipo 078 (RT078) ha adquirido un grupo de 4 genes implicados en el metabolismo de la trehalosa. Entre ellos, una permeasa de tipo PTS (fosfotransferasa) es, al mismo tiempo, necesaria y suficiente para el crecimiento de las cepas, incluso con bajas concentraciones de trehalosa. Los autores plantean que la aplicación de la trehalosa como aditivo alimentario en la alimentación humana, poco tiempo antes del surgimiento de estos dos ribotipos epidémicos, ha contribuido a su aparición y a su hipervirulencia [1].
¿Qué sabemos sobre este tema?
El análisis por secuenciación del genoma completo de cepas de C. difficile ribotipo 027 (RT027) ha demostrado que dos linajes independientes han surgido en América del Norte entre el 2000 y el 2003 [2]. La comparación con cepas RT027 históricas previas a la epidemia ha evidenciado que los linajes epidémicos han adquirido una mutación en el gen gyrA, que ha conducido a una mayor resistencia a los antibióticos del grupo de las fluoroquinolonas. Aun cuando el desarrollo de esta resistencia ha desempeñado ciertamente un rol en la propagación de las cepas RT027, también se ha observado en ribotipos de C. difficile no epidémicos e identificados en cepas que datan de mediados de la década del 80. Por consiguiente, es probable que otros factores hayan contribuido a la emergencia de cepas epidémicas RT027. La prevalencia de un segundo ribotipo de C. difficile, RT078, se multiplicó en los hospitales y las clínicas entre 1995 y 2007 y se asoció con una mayor severidad [3]. No obstante, los mecanismos involucrados en el aumento de la virulencia siguen siendo desconocidos. Dado que los linajes RT027 y RT078 están filogenéticamente alejados uno de otro, es posible que los cambios que condujeron en forma simultánea a un aumento de la prevalencia y de la gravedad de la infección puedan provenir de mecanismos independientes.
¿Cuáles son los principales resultados que aporta este estudio?
Se ha demostrado que las cepas del ribotipo RT027 tienen una ventaja competitiva comparada con las otras cepas in vitro y en modelos murinos de infección con C. difficile. Para investigar los mecanismos involucrados, los autores han examinado el uso de diferentes fuentes de carbono por las diversas cepas y han evidenciado una mayor capacidad para utilizar el disacárido trehalosa en el caso de las cepas del ribotipo RT027. Comparando los genomas de numerosas cepas de C. difficile, los autores han identificado una potencial enzima fosfotrehalasa (TreA) que metaboliza la trehalosa-6-fosfato en glucosa y glucosa-6-fosfato. Luego, los autores observaron que las cepas del ribotipo RT027 activaban ese gen con una concentración 500 veces menor de trehalosa que las otras cepas de C. difficile. Análisis más finos permitieron identificar un polimorfismo en el represor transcripcional (TreR) de TreA en todas las cepas del ribotipo RT027 y en otras cepas cercanas al origen epidémico en Europa y Australia. Para evaluar si la capacidad de metabolizar la trehalosa tenía un impacto en la virulencia, los autores la administraron a ratones trasplantados con una microbiota humana e infectados por una cepa del ribotipo RT027 (R20291), o bien infectados por la misma cepa, pero delectada para el gen TreA (R20291Δ TreA) y, por ende, incapaz de metabolizar la trehalosa. La mortalidad era mucho menor con esta última cepa R20291Δ TreA (Figura 1).
Imagen
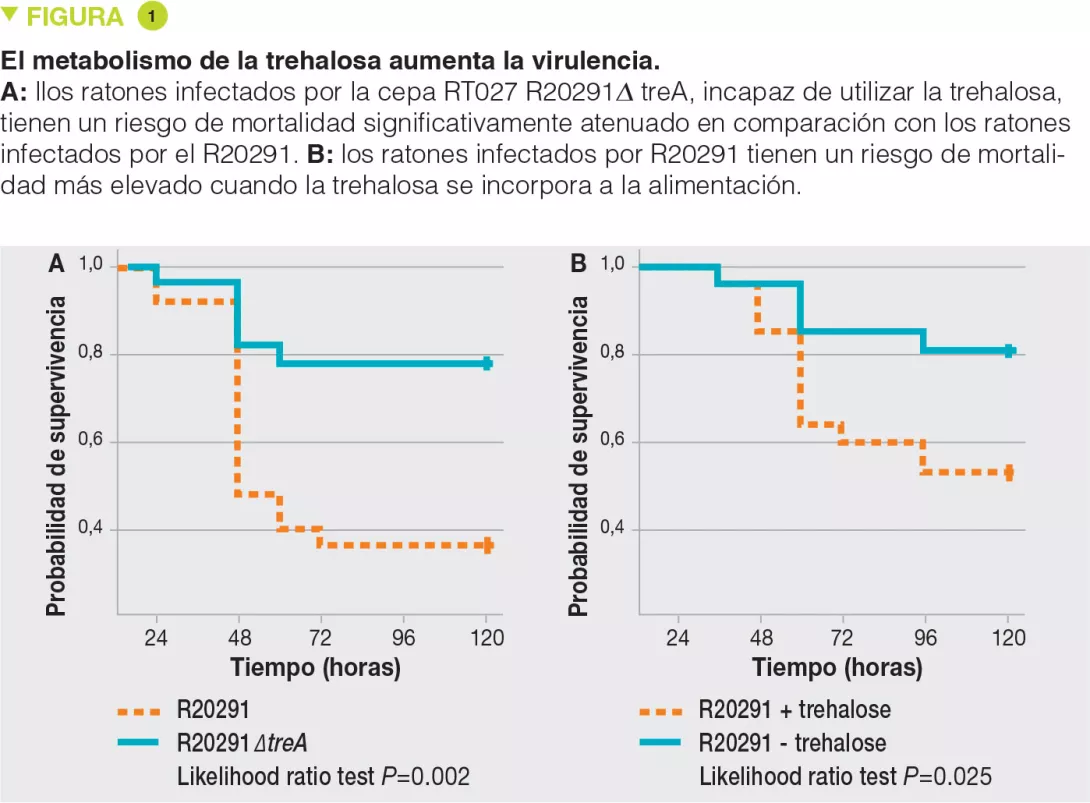
En una segunda experiencia, los autores infectaron ratones con una microbiota humana con la cepa del ribotipo RT027 (R20291) en presencia o en ausencia de trehalosa en el agua para beber (suministrada en una dosis equivalente a la recibida en una comida humana). La mortalidad era muy superior en presencia de trehalosa. Las dos experiencias acumuladas confirman la hipótesis según la cual la trehalosa en los alimentos contribuye a la severidad de las cepas del ribotipo RT027. El análisis genético de las cepas del ribotipo RT078 ha mostrado la inserción de 4 genes que codifican una 2.a copia de la fosfotrehalasa (TreA2) y de su represesor (TreR2) y otros 2 genes vinculados. Un enfoque por mutación y sobreexpresión ha permitido confirmar que esta inserción era responsable de la capacidad de las cepas del ribotipo RT078 de desarrollarse en presencia de trehalosa.
¿Cuáles son las consecuencias en la práctica?
La trehalosa es un azúcar extremadamente estable, resistente tanto a las altas temperaturas como a la hidrólisis. Considerada ideal para un uso en la industria agroalimentaria, su empleo no se propagó hasta el 2000, fecha en que se descubrió un nuevo procedimiento de producción a bajo costo [3]. Su uso ha sido autorizado en la alimentación por la Administración de Alimentos y Medicamentos (FDA) estadounidense en el año 2000 y por las instituciones europeas en el año 2001. La amplia adopción de la trehalosa coincide con la aparición de los focos de infección a causa de las cepas RT027 y RT078. El conjunto de estos resultados sugieren un rol causal de la trehalosa incorporada a los alimentos en la aparición de estas cepas epidémicas hipervirulentas de C. difficile.